

Sci. Pharm. 2024, 92(2), 21; https://doi.org/10.3390/scipharm92020021 - 17 Apr 2024
Abstract
Antimicrobial resistance threatens the effective prevention and treatment of an increasingly broad spectrum of infections caused by pathogenic microorganisms. This pressing challenge has intensified the search for alternative antibiotics with new pharmacological properties. Due to the chemical synergy between the biological activity of
[...] Read more.
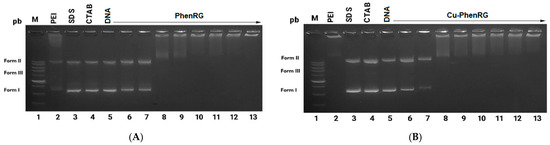
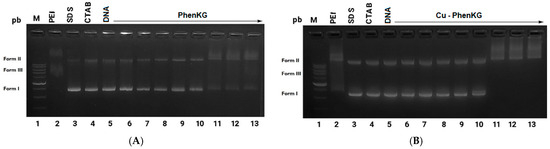
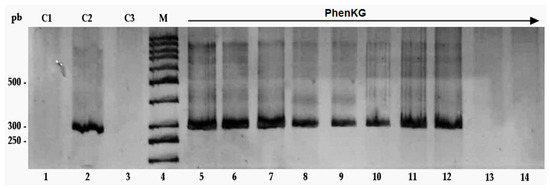
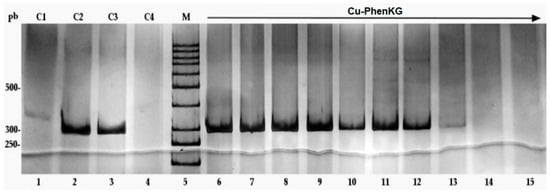
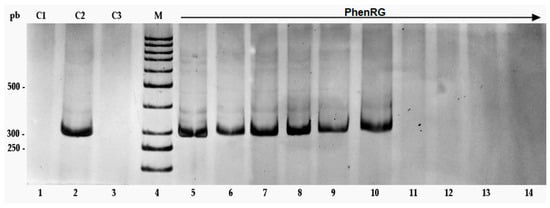
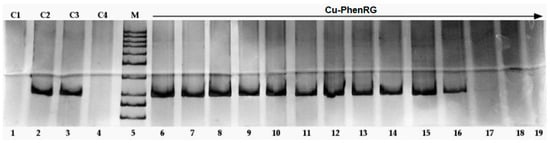
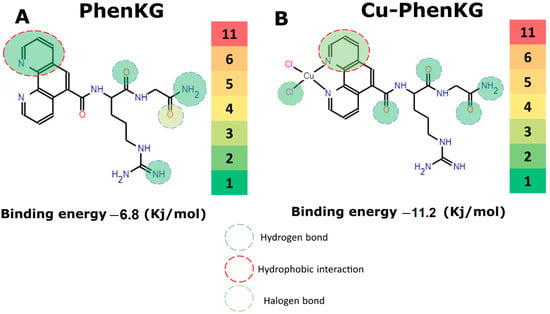
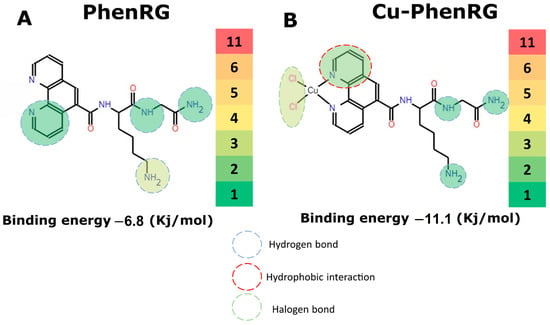
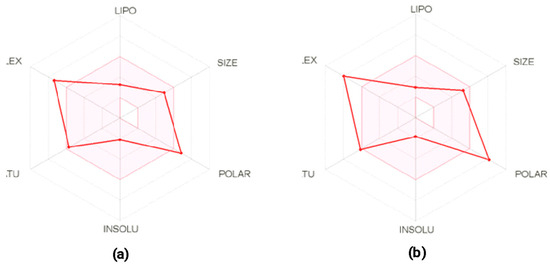
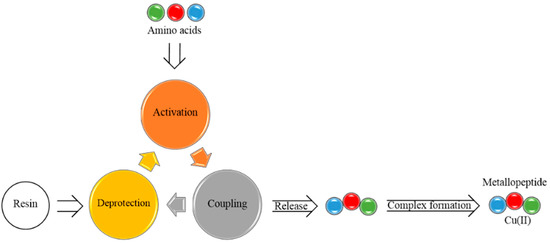
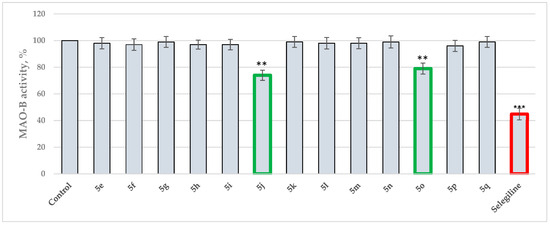
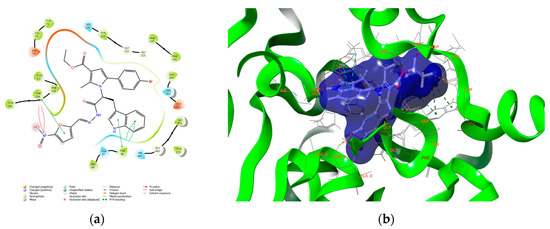
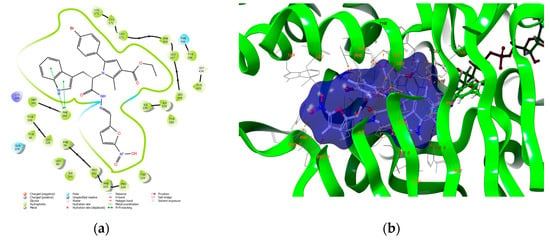
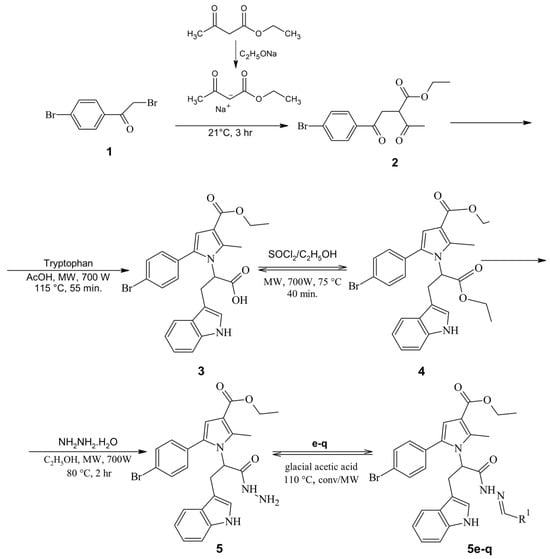
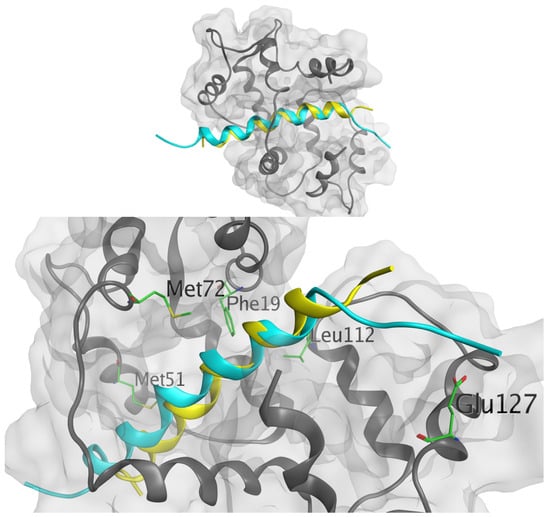
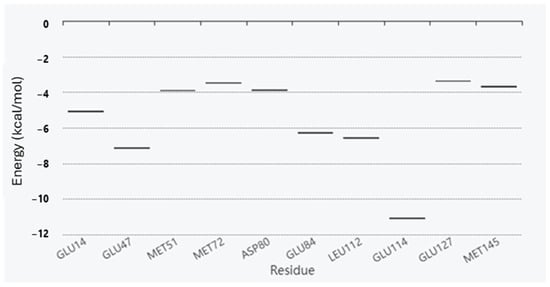
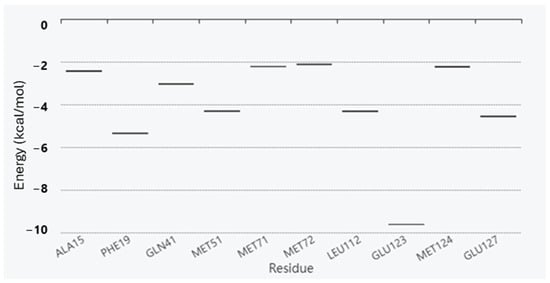
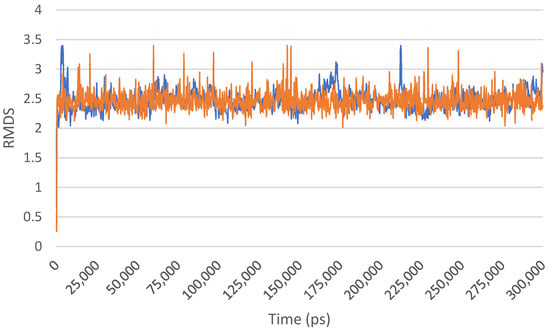
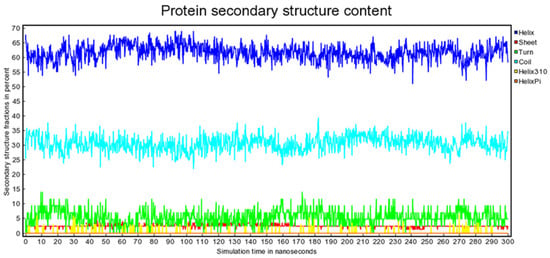
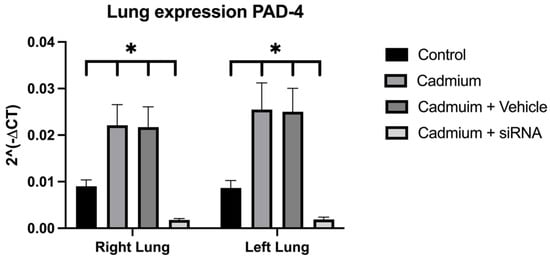
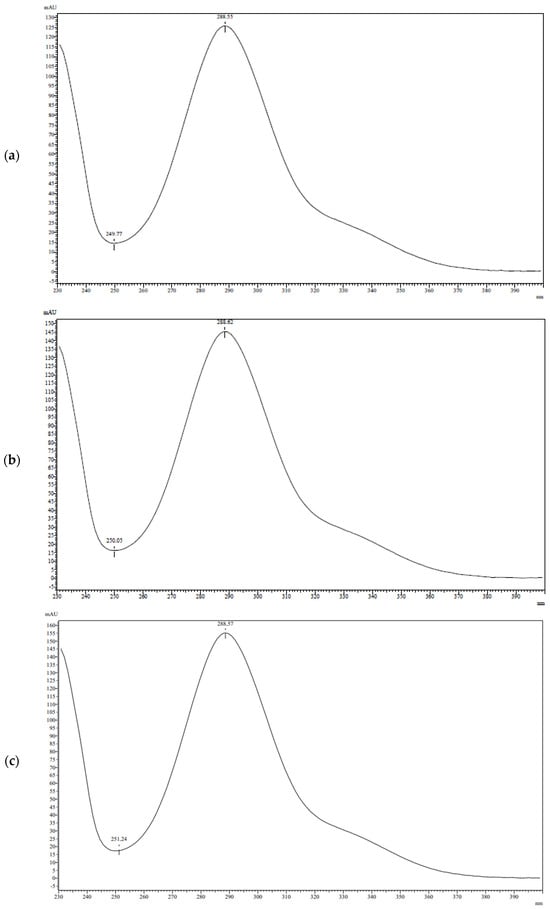
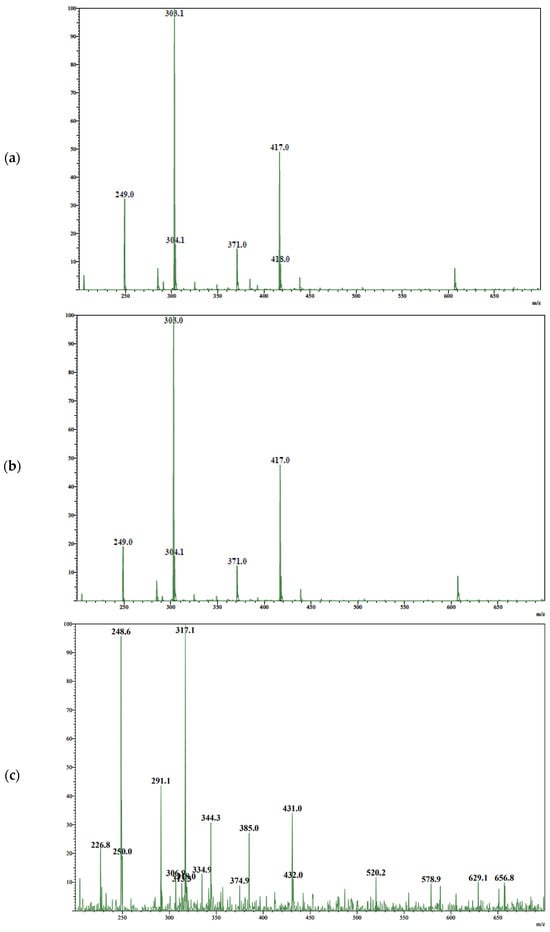
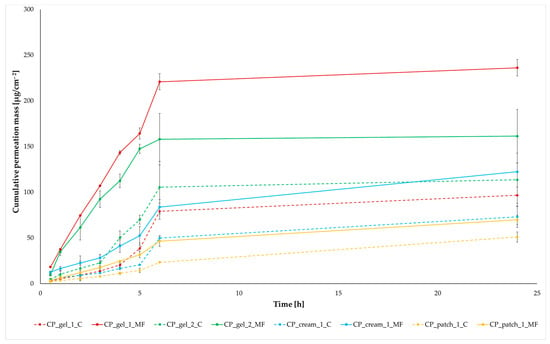
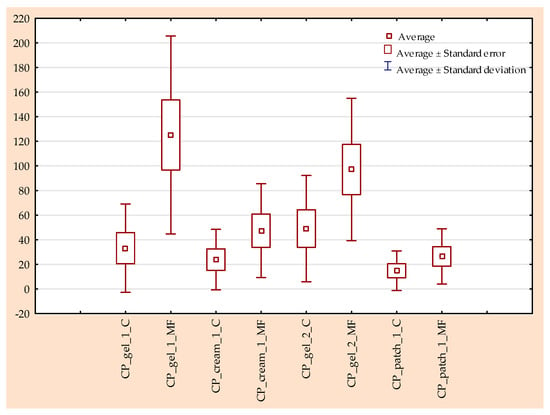
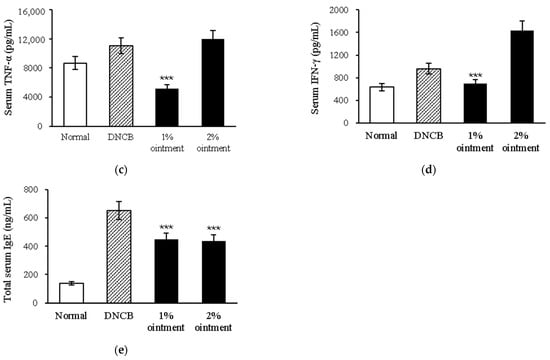
Antimicrobial resistance threatens the effective prevention and treatment of an increasingly broad spectrum of infections caused by pathogenic microorganisms. This pressing challenge has intensified the search for alternative antibiotics with new pharmacological properties. Due to the chemical synergy between the biological activity of antimicrobial peptides (AMPs) and the different modes of action, catalytic properties, and redox chemistry of metal complexes, metallopeptides have emerged in recent years as an alternative to conventional antibiotics. In the present investigation, peptide ligands conjugated with 5-carboxy-1,10-phenanthroline (Phen) were prepared by solid-phase peptide synthesis (SPPS), and the corresponding copper(II) metallopeptides, Cu-PhenKG and Cu-PhenRG (where K = lysine, R = arginine, and G = glycine), were synthesized and characterized. The antimicrobial activities of these compounds toward Gram-positive and Gram-negative bacteria, evaluated by the broth microdilution technique, indicate that the metal center in the metallopeptides increases the antimicrobial activity of the complexes against the conjugated peptide ligands. Minimum inhibitory concentration (MIC) values of 0.5 μg/mL for S. aureus with the Cu-PhenKG complex and 0.63 μg/mL for S. typhimurium with the Cu-PhenRG complex were obtained. The MIC values found for the conjugated peptides in all microorganisms tested were greater than 1.5 μg/mL. The interactions of the conjugated peptides and their metallopeptides with plasmid DNA were evaluated by agarose gel electrophoresis. Alterations on the replication machinery were also studied by polymerase chain reaction (PCR). The results indicate that the complexes interact efficiently with pBR322 DNA from E. coli, delaying the band shift. Furthermore, the resulting DNA–metallopeptide complex is not a useful template DNA because it inhibits PCR, since no PCR product was detected. Finally, molecular dynamics and molecular docking simulations were performed to better understand the interactions of the obtained compounds with DNA. The Cu-PhenRG complex shows a significantly higher number of polar interactions with DNA, suggesting a higher binding affinity with the biopolymer.
Full article
(This article belongs to the Special Issue Feature Papers in Scientia Pharmaceutica)
►
Show Figures
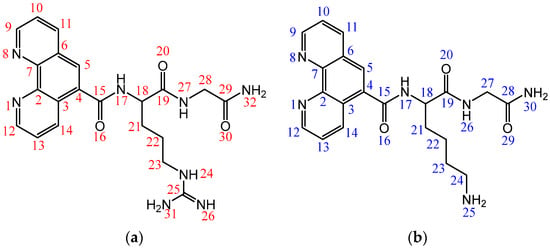
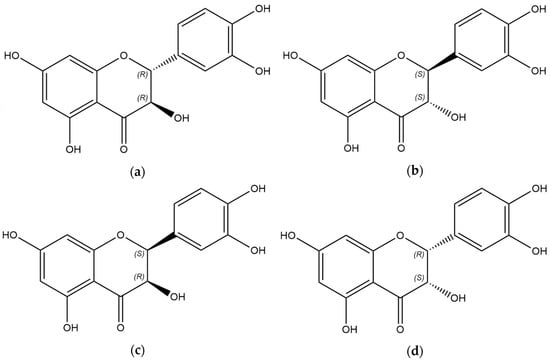
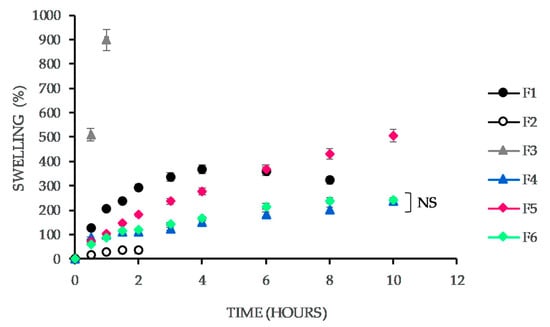
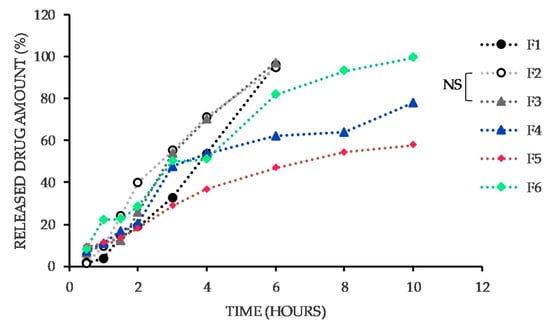
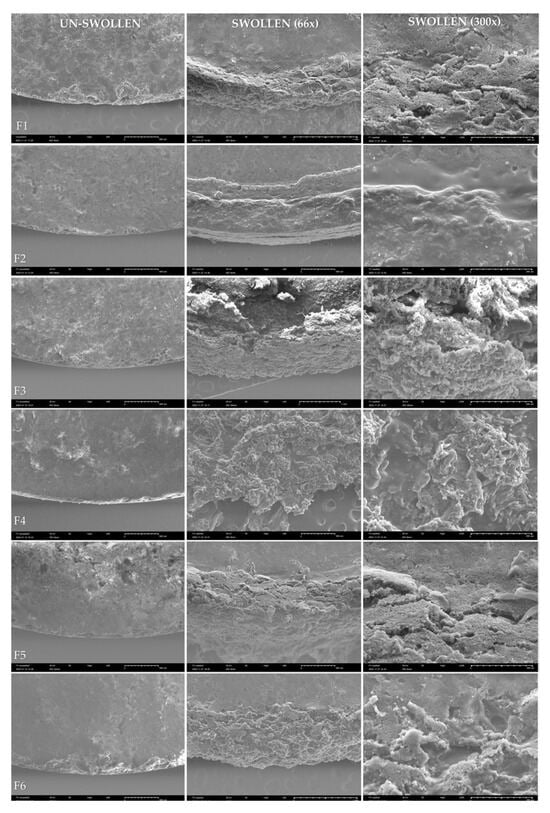
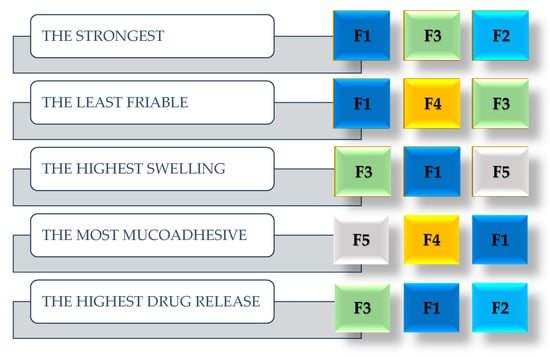
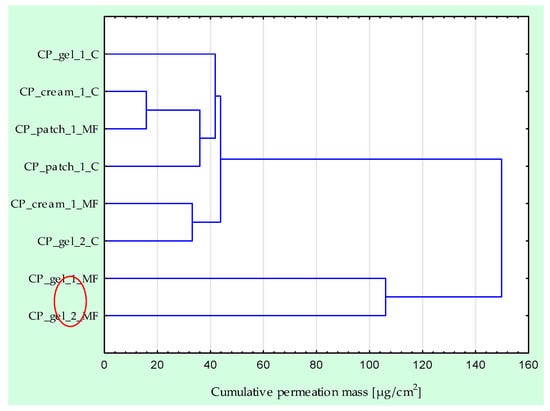
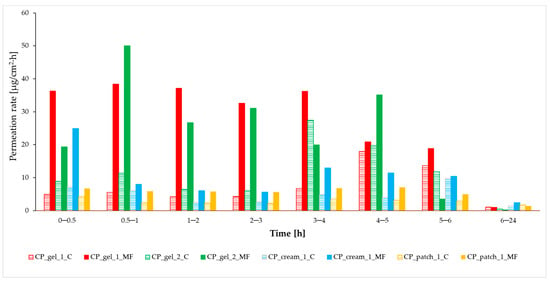
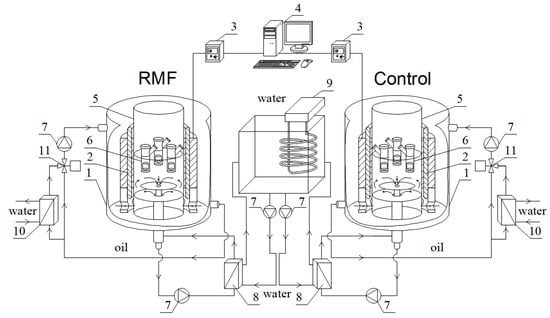
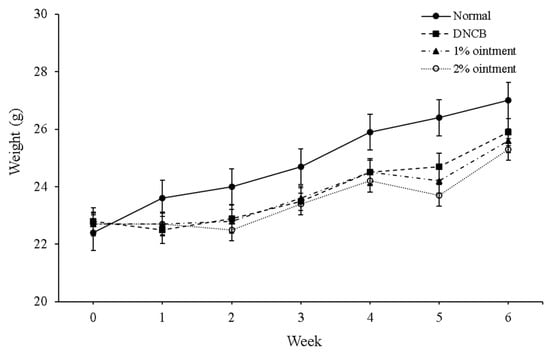
Figure 1




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}